Rapid Quantification of Trypsin Inhibitors in Food and Feed Formulations with Electrospray Ionization Mass Spectrometry
Special Issues
A rapid, accurate, and precise method for the quantification of trypsin inhibitor activity was evaluated. The method utilizes alpha hydroxyl acid capped oligo-lysines [hydroxy acid (Lys)n] or alpha hydroxyl acid capped oligo-lysines-methionine [hydroxy acid (Lys-Met)] as substrates. Hydrolysis of the oligopeptides yields unique chemical residues that were readily quantified with electrospray–mass spectrometry (ESI-MS). Accuracy and precision of the approach compared favorably with that of the standard test method.
A rapid, accurate, and precise method for the quantification of trypsin inhibitor activity was evaluated. The method uses alpha-hydroxyl acid–capped oligolysine (hydroxy acid [Lys]n) or alpha-hydroxyl acid–capped oligolysine-methionine (hydroxy acid [Lys-Met]) as substrates. Hydrolysis of the oligopeptides yields unique chemical residues that were readily quantified with electrospray ionization mass spectrometry (ESI-MS). Accuracy and precision of the approach compared favorably with that of the standard test method.
The nutritional importance of legumes as an economic source of proteins is well recognized (1). Legumes provide a good balance of proteins, amino acids, and essential lipids, resulting in high overall nutritional value. Legumes are the leading source of protein in human diets in many countries and they are also important sources of protein in animal feed (2). However, legumes also contain many antinutritional factors (ANFs), the principal one being the trypsin inhibitors, which are a serious obstacle to full utilization of legumes in human diets and animal feed because these proteins deactivate (inhibit) trypsin catalyzed hydrolysis of proteins and peptides (3). Trypsin inhibitors are widespread and are present in many plants and some animal species. They have been reported in various legumes such as soybeans (4), lima beans (5), navy beans (6), black eyed peas (7), and Jatropha curcas (8). Trypsin is a serine protease that breaks down proteins in the digestive tract of animals. Trypsin cleaves peptide chains at the carboxyl side of the basic amino acids lysine and arginine, except when either is followed by proline (9). Thus, the enzyme plays an important role in the digestion of proteins and uptake of amino acids. Trypsin inhibitors manifest their action in two ways, either by blocking the substrate from the enzyme active site or by modifying the amino acid residues of the enzyme, thus reducing its activity. The end result is decreased utilization of proteins, thereby reducing the nutritional value of grain and legumes. To counteract these effects, processing of the seeds before ingestion is often recommended and widely practiced. Heat treatment is perhaps the most commonly used and effective method for eliminating or reducing trypsin inhibitor content. Traditional heat treatment processing includes dehulling, soaking, and heating (10). The use of moderate heat treatment causes the partial denaturation of proteins and generally has a beneficial effect on the nutritional value; by facilitating enzyme access it makes proteins more digestible (11,12). Measuring the trypsin inhibitors in legumes is of importance to the food and feed processors, who are concerned with providing a high-quality product for animal feed. Therefore, it is important to do accurate measurements for the trypsin inhibitor content in the food and feed formulations.
Classical Methods for the Quantification of Trypsin Inhibitors
At present the trypsin inhibitor content of soy products is determined through the standard American Association of Cereal Chemists (AACC) method 22-40.01 or a similar method such as International Organization for Standardization (ISO) 14902 (13,14). The AACC method is based on monitoring the decrease in trypsin induced hydrolysis of a synthetic substrate N-α-benzoyl-DL-arginine-p-nitroanilide hydrochloride (BAPA) in the presence of trypsin inhibitors. BAPA hydrolysis leads to the release of p-nitroaniline, a chromophore that absorbs radiation in the blue region at λ410 nm. BAPA hydrolysis is depicted in Figure 1.


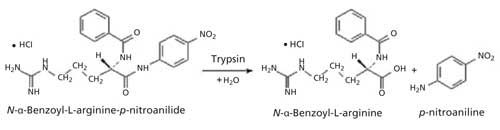
Figure 1: Trypsin-catalyzed hydrolysis of BAPA.
The presence of trypsin inhibitors in the aqueous reaction medium leads to “poisoning” of trypsin, which leads to retardation or complete cessation of enzyme activity, a reduced release of p-nitroaniline, and lower absorption at λ410 nm. The amount of p-nitroaniline formed during a specified time period is measured spectrophotometrically at λ410 nm. The original procedure was reported by Kakade and colleagues and adopted as the AACC standard method 71-10 (15,16). Further modifications in the method were suggested by Hamerstrand and colleagues that were incorporated into the current AACC standard method 22-40.01 (17).
However, the modified method still suffers from problems stemming from inherent nonlinearity of absorption, which can affect accuracy and precision unless the procedure is adhered to within a very restrictive regime (18). Discrepancies in results obtained with two BAPA-based standard methods ISO 14902 and AACC 22-40.01 were reported recently by Sueiro and colleagues (19). The ISO 14902 method differs from AACC 22-40.0 primarily in the choice of the substrate. The ISO method requires the use of enantiomerically pure N-α-benzoyl-L-arginine-p-nitroanilide, whereas the AACC method recommends the use of racemic N-α-benzoyl-DL-arginine-p-nitroanilide. Thus, both methods are subject to nonlinearity issues related to absorption-based determinations. To overcome this shortcoming of the spectrophotometry based methods, an electrospray ionization mass spectrometry (ESI-MS)-based approach was evaluated. The approach relies on lysine containing a synthetic substrate, whose concentration in feed suspensions can be readily monitored with ESI-MS. The approach is immune to inherent nonlinearity of absorption-based methods and should yield accurate and precise quantification of trypsin inhibitors in food and feed formulations.
The method relies on trypsin-catalyzed hydrolysis of the synthetic substrate hydroxy methyl thiobutyric acid–capped dilysine (HMTBA-[Lys]2), which was synthesized through a papain-catalyzed reaction with lysine-ethyl ester (Lys-EE) as the substrate. The HMTBA-(Lys)2 was synthesized through a chymotrypsin-catalyzed reaction of dilysine and HMTBA-ethyl ester (20). It was hypothesized that HMTBA-(Lys)n species should be suitable substrates for monitoring trypsin activity because trypsin is known to cleave peptide bonds at the carboxylic end of basic amino acids arginine and lysine unless the amino acid down side of the arginine or lysine is proline. Hydrolysis of HMTBA-(Lys)n results in the release of a free lysine leading to a decrease in the concentration of the selected HMTBA-(Lys)n substrate. HMTBA-(Lys)n yields characteristic ions in ESI-MS and their concentrations can be readily monitored. Trypsin activity can be determined by monitoring the extent hydrolysis of suitable substrates such as HMTBA-(Lys)2 (Figure 2).


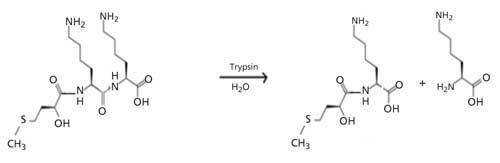
Figure 2: Trypsin-catalyzed hydrolysis of HMTBA-(Lys)2.
A decrease in the rate of HMTBA-(Lys)2 hydrolysis in the presence of trypsin inhibitors extracted from soybean meal and other plant materials with ESI-MS can be used to assess the concentration of trypsin inhibitors in plant materials, thus offering an approach that is less susceptible to the nonlinearity issue.
Experimental
The overall experimental approach involved the following steps:
- Synthesis of HMTBA-(Lys)2.
- Optimization of trypsin-catalyzed hydrolysis of HMTBA-(Lys)2 and monitoring of trypsin-catalyzed hydrolysis with ESI-MS.
- Extraction of trypsin inhibitors from soybean and Jatropha meals.
- Monitoring of trypsin inhibitor activity in soybean and Jatropha meal samples with AACC 22-40.01 and with ESI-MS with HMTBA-(Lys)2 as the substrate.
Synthesis of Dilysine
Dilysine was synthesized through a papain-catalyzed reaction with lysine ethyl ester as the substrates in a three phase reaction media, comprised of water, n-octane (C8H18), and decafluoropentane (C5H2F10), the incubation temperature was set at 37 °C. The number of lysine residues in the oligomer depends on the incubation period. The predominant product after a 2-h incubation period was found to be dilysine. The product yield was monitored with ion-pair liquid chromatography (LC) and ESI-MS.
Characterization of Dilysine
Dilysine prepared through papain catalyzed reaction was characterized with ESI-MS and ion-pair chromatography on a C18 column.
Synthesis of HMTBA-(Lys)2
HMTBA-capped peptides were synthesized through a chymotrypsin catalyzed reaction in phosphate buffer. Dilysine and HMTBA ethyl ester were added to a reaction flask containing 50 mM sodium phosphate (dibasic) buffer (pH 7.8). An enzyme suspension (3.33 mg/mL of enzyme in dibasic sodium phosphate buffer) was added to the flask. The reaction mixture was incubated at 37 °C in an incubator–shaker for 180 min. After the reaction an aliquot was taken for analysis. The aliquot was diluted proportionately, centrifuged, and filtered, then 5 µL of the filtered product was introduced into the ESI-MS system for determining the product profile.
Preparation of Reagents for Determining Trypsin Inhibitors
Reagents for trypsin inhibitor determinations with HMTBA-(Lys)2 as the substrate were prepared in a manner similar to that described in the AACC 22-40.01 standard method for determination of trypsin inhibitors in soybean meal:
- Sodium phosphate buffer, pH 7.6, at 25 °C (1.67 mM sodium phosphate, pH was adjusted with 1 N sodium hydroxide).
- Substrate HMTBA-(Lys)2 1.0 mM, prepared by dissolving 406 mg of HMTBA-(Lys)2 in 1 L of water.
- Trypsin solution was prepared by dissolving 3.0 mg of trypsin (Sigma-Aldrich T9201) in 200 mL of 1 mM hydrochloric acid. The solution was stored in a refrigerator at 4 °C before use.
- Soybean and Jatropha meal were used for assessing efficacy of the method. Untreated meals were obtained by grinding raw beans and passing meals through 100-mesh screen. Treated meals were obtained through wet heat treatment of raw screen meals at 130 °C. Next, 1-g portions of meals were stirred with 50 mL of 0.01 N sodium hydroxide for 3 h at 25 °C; the pH of the extract was maintained at 8.5–10.0. The meal suspensions were transferred to 100-mL volumetric flasks and the volume was brought to the mark with high performance liquid chromatography (HPLC)-grade water.
- BAPA solution was in accordance with the standard method AACC 22-40.01.
Sample-Free Enzymatic Hydrolysis of HMTBA-(Lys)2
Hydrolysis of the substrate was investigated before the initiation of the inhibition assay. First, 5-mL aliquots of HMTBA-(Lys)2 solution were taken and introduced into 15-mL vials. Then 3 mL of HPLC-grade water was added to each vial. Vials were placed in an incubator–shaker maintained at 37 °C for 30 min. After equilibration, 2 mL of trypsin solution, equilibrated at 37 °C, was added to each vial. Vials were removed from the incubator shaker after set equilibration periods (0–20 min) and the enzyme was heat deactivated. Aliquots were drawn from the vials and filtered through 0.2-µm filters, and a 5-µL portion of the filtered solution was introduced into the ESI-MS system. Intensity of the protonated ion of HMTBA-(Lys)2 at m/z 407 was monitored.
Substrate-Free Enzymatic Reaction Blanks
First, 7 mL of HPLC-grade water along with 1 mL of soybean meal extract was added to 15-mL vials. The vials were placed in an incubator–shaker maintained at 37 °C for 30 min. After equilibration, 2 mL of trypsin solution was added to each vial. The vials were removed from the incubator shaker after 10-min equilibration periods and the enzyme was deactivated. The contents of the vials were allowed to cool to 25 °C. Aliquots were drawn from the vials and filtered through 0.2-µm filters, and a 5-µL portion of the filtered solution was introduced into the ESI-MS system.
Deactivated Enzyme Samples
Enzyme blank samples were prepared by adding 2 mL of trypsin solution to 15-mL vials. The enzyme solutions were heated to 85 °C for 10 min. Solutions were allowed to cool down to 25 °C. Then 5-mL aliquots of HMTBA-(Lys)2 were taken and introduced into vials. Next 2 mL of HPLC-grade water along with 1 mL of soybean meal or Jatropha meal extract was added to the vials. The vials were placed in an incubator–shaker and equilibrated at 37 °C. The contents were allowed to react for 10 min. The contents of the vials were cooled down to room temperature, aliquots were filtered through a 0.2-μm filter, and 5-μL aliquots of the filtered solutions were introduced into the ESI-MS system.
Measurement of Trypsin Inhibitor Activity in Soybean and Jatropha Meal Extracts
Soybean and Jatropha meal extracts were mixed with HPLC-grade water in varied ratios, 1 mL of the undiluted or the diluted extracts were mixed with 5 mL of HMTBA-(Lys)2 solution and 2 mL of HPLC-grade water. The contents of the vials were mixed, then 2 mL of trypsin solution was added to each vial. The vials were placed in the incubator shaker maintained at 37 °C and incubated for 10 min. The vials were transferred to a water bath maintained at 85 °C to terminate the reaction. The contents of the vials were allowed to cool down to 25 °C. Aliquots were drawn from vials and filtered through 0.2-µm filters. A 5-µL aliquot of the filtered solutions was introduced into the ESI-MS system.
ESI-MS Parameters
Quantification of HMBTA-(Lys)2 was carried out through direct infusion of 5-μL sample aliquots through a fixed volume loop. Samples were carried into the Varian/Agilent 320 ESI-MS system with acetonitrile-water mixture at 220 μL/min. The ESI-MS system was operated in the positive ion mode. Operation parameters were as follows:
Needle voltage: 4500 V
Shield voltage: 600 V
Capillary voltage: 80 V
Drying gas temperature: 200 °C
Nebulizing gas: nitrogen
Drying gas: nitrogen
Results and Discussion
Characterization of HMTBA-(Lys)2
The purity of HMTBA-(Lys)2 used as the substrate was established through reversed-phase LC and ESI-MS. Reversed-phase LC of the HMTBA-(Lys)2 preparation showed the presence of one dominant peak that accounted for >95% of the area of the total peak areas in the chromatogram. The ESI-MS spectrum of the HMTBA-(Lys)2 solution showed a dominant ion at m/z 407, corresponding to [HMBTA-(Lys)2 + H]+ (Figure 3).


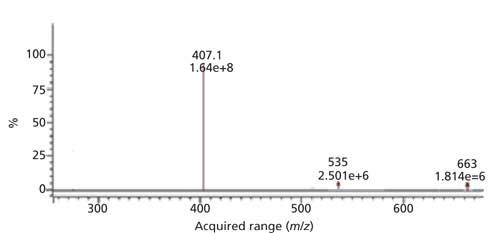
Figure 3: ESI-MS positive ion spectrum of HMTBA-(Lys)2.
The ions at m/z 535 and 663 correspond to HMTBA-(Lys)3 and HMTBA-(Lys)4 were observed, however, their combined concentration was less than 5%. The linearity of ESI-MS response for HMTBA-(Lys)2 was monitored as intensity of m/z 407 over concentrations of 1–203 μg/mL in aqueous solutions containing soybean or Jatropha meal extracts, response was found to be linear with a correlation coefficient of 0.998–1.002.
Trypsin-Catalyzed HMTBA-(Lys)2 Hydrolysis
Trypsin-catalyzed hydrolysis of HMTBA-(Lys)2 was monitored over incubation periods varying from 0 to 20 min. The extent of hydrolysis was monitored by measuring the intensity of protonated HMTBA-(Lys)2 ion at m/z 407 and its hydrolysis product HMTBA-Lys (MW 278), which appears at m/z 279 with ESI-MS. Hydrolysis led to the appearance of protonated HMTBA-Lys ion at 279 m/z. The appearance of the latter ion can provide an independent means to monitor the extent of trypsin-mediated hydrolysis of the substrate; however, it was not examined during this study. Results showed that HMTBA-capped dilysine is rapidly catalyzed by trypsin and therefore a good substrate for monitoring trypsin-catalyzed hydrolysis. The change in HMTBA-(Lys)2 concentration with the incubation period is shown in Figure 4. As expected, the enzymatic hydrolysis of HMTBA-(Lys)2 occurs at a considerably faster rate than that reported for BAPA, perhaps because of its higher solubility in water and also because it is a more natural substrate for trypsin. Results showed that HMTBA-(Lys)2 is a suitable substrate for monitoring trypsin-catalyzed hydrolysis of the amide bond at the carboxylic end of lysine.


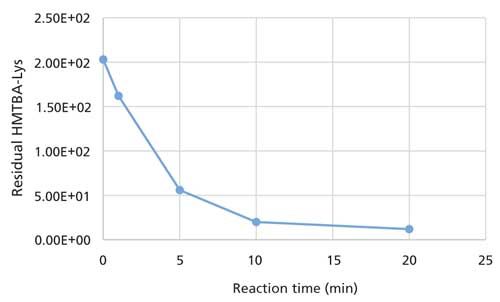
Figure 4: Change in HMTBA-(Lys)2 concentration with reaction time.
Substrate-Free Enzymatic Reactions
Filtered aliquots from reactions carried with soybean and Jatropha meal extracts in the presence of trypsin but without HMTBA-(Lys)2 were introduced into the ESI-MS system. ESI-MS mass spectra for the reaction media aliquots were acquired in both the scan over m/z 200–450 and SIM mode set for monitoring ion m/z 407. The scan mode spectra showed the presence of characteristic background ions observed with acetonitrile-water mixtures at very low intensities. Ions at m/z 407 were not observed at all or observed at intensities three orders of magnitude lower than ones observed in the presence of the HMTBA-(Lys)2. These results support the contention that HMTBA-(Lys)2 is a suitable substrate for monitoring trypsin catalyzed hydrolysis.
Substrate Concentration in Reaction Media with Deactivated Enzyme
Filtered aliquots from reaction media incubated at 37 °C for 10 min with deactivated trypsin containing HMTBA-(Lys)2 and soybean and Jatropha meal extracts were introduced into the ESI-MS system. ESI-MS mass spectra for the reaction media aliquots were acquired in the selected-ion monitoring (SIM) mode set for ions m/z 407 and m/z 279. Only the ion at m/z 407 was observed in the reaction with either the soybean or Jatropha meal extracts. Results showed that HMTBA-(Lys)2 concentration did not change after incubation with the deactivated enzyme. Integration results for m/z 407 in reaction media before and after incubation are shown in Figure 5.


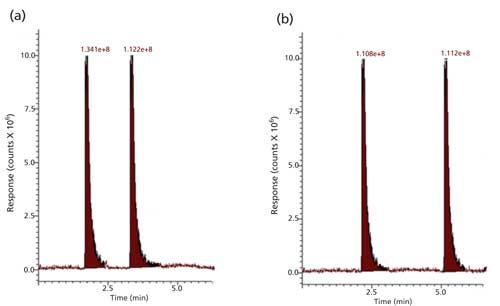
Figure 5: (a) Intensity of HMTBA-(Lys)2 ion m/z 407 in reaction media at the start of incubation with deactivated trypsin. (b) Intensity of HMTBA-(Lys)2 ion m/z 407 in reaction media after a 10-min incubation with deactivated trypsin.
Measurement of Trypsin Inhibition
Analysis for substrate HMBTA-(Lys)2, in reaction media containing the substrate, trypsin with soybean or Jatropha meal extracts showed that the intensity of protonated HMTBA-(Lys)2 ion at m/z 407 in the reaction media was related to the amount of meal extracts introduced, intensity of m/z 407, that is, the intact (unhydrolyzed) HMTBA-(Lys)2 increased indicating a decrease in the extent of HMTBA-(Lys)2 hydrolysis through trypsin inhibition. The increase in intensity of ion m/z 407 relative to reaction media without the soybean or Jatropha extracts was related to the volumes of meal extracts introduced into the reaction media. It was observed that meal extracts of untreated (raw) soybean and Jatropha meal were more effective in inhibiting the hydrolysis than the extracts of the heated treated meals. Extracted ion chromatograms for m/z 407 obtained for three replicate injections of reaction media aliquots after a 10-min incubation with trypsin in the absence of soybean or Jatropha meal extract are shown in Figure 6. Extracted ion chromatograms for m/z 407 obtained for three replicate injections of reaction media aliquots after a 10-min incubation with trypsin in the presence of diluted Jatropha meal extract (twofold dilution) are shown in Figure 7.


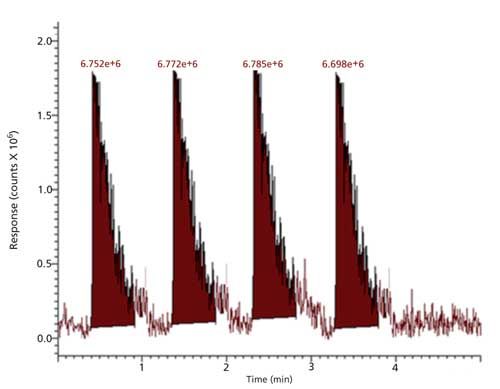
Figure 6: Intensity of HMTBA-(Lys)2 ion m/z 407 in reaction media after a 10-min incubation with active trypsin.


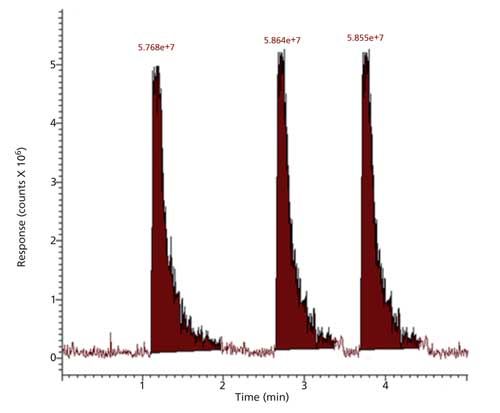
Figure 7: Intensity of HMTBA-(Lys)2 ion m/z 407 in reaction media after a 10-min incubation with trypsin and Jatropha meal extract.
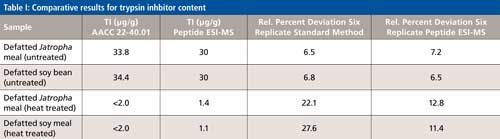
Results show the addition of diluted Jatropha meal reduced HMTBA-(Lys)2 to half the value obtained in the absence of the extract. In the standard BAPA-based trypsin inhibitor assay a linear relationship between BAPA hydrolysis over a set time (10 min) is assumed. If a similar relationship is assumed in the present case this would imply that diluted Jatropha deactivates half of the enzyme (~15 μg) present in the reaction media. Thus, trypsin inhibitor content of the meal would be 30 μg/g, a value similar to the one obtained with the AACC standard method. Results obtained for untreated and treated soybean and Jatropha meals with the two methods are shown in Table I.


CLICK TABLE TO ENLARGE
Conclusions
The results of this short set of experiments clearly demonstrate that rapid, specific and accurate determination of trypsin inhibitors is feasible with ESI-MS, the approach does not suffer from nonlinearity issues, which mandate careful control of all experimental parameters in absorption based determinations. However, collaborative multilaboratory efforts are required before the ESI-MS-based approach can be fully validated.
References
- M. Borget, Food Legumes (Macmillan, London, 1992).
- G.P. Gent, Cur. Plant Sci. Biotechnol. Agric.19, 361–399 (1994).
- K.A. Wilson, Antinutrients and Natural Toxicants in Foods, Proceedings of a 1979 symposium, 187–202 (1981).
- W.E. Ham and R.M.J. Sandstedt, Biol. Chem.154, 505–506 (1944).
- H. Fraenkel-Conrat, R.C. Bean, E.D. Ducay, and H.S. Olcott, Arch. Biochem. Biophys.37, 393–407 (1952).
- L.P. Wagner and J.P. Riehm, Arch. Biochem. Biophys.121, 672–677 (1967).
- M.M. Ventura and J.X. Filho, Acad. Bras. Cienc. Anais.38, 553–557 (1966).
- H.P.S. Makkar, K. Becker, and M. Wink, J. Agric. Food Chem.45, 3152–3157 (1997).
- J.V. Olsen, S.E. Ong, and M. Mann, Molecular & Cellular Proteomics3(6), 608–614 (2004).
- J. Haddad and K. Allaf, J. Food Engineering79, 353–357 (2007).
- M.C. Marquez and R. Alonso, J. Food Composition and Analysis12, 211–217 (1999).
- American Association of Cereal Chemists, Approved Methods of Analysis, 11th Ed., “Method 22-40.01. Measurement of Trypsin Inhibitor Activity of Soy Products - Spectrophotometric Method” (first approved Nov. 7, 1973; reapproved Nov. 3, 1999, AACC International, St. Paul, Minnesota, Doi: 10. 1094/AACC Int. Method-22.40.01).
- International Organization for Standardization, Standard 14902:2001. Animal Feedstuffs - Determination of Trypsin Inhibitor Activity of Soya Products (approved Oct. 2001; reapproved Aug. 2012, International Organization for Standardization, Geneva, Switzerland).
- M.L. Kakade, N. Simons, and I.E. Liener, Cereal Chem.46, 518–526 (1969).
- M.L. Kakade, J.J. Rackis, J.E. Mcghee, and G. Puski, Cereal Chem.51, 376–381 (1974).
- G.E. Hamerstrand, L.T. Black, and J.D. Glover, Cereal Chem.58, 42–45 (1981).
- C. Stauffer, Cereal Chem.67, 296–302 (1990).
- S. Sueiro, M. Hermida, M. Gonzalez, A. Lois, and J.L. Rodriguez-Otero, J. Am. Oil Chem. Soc.92, 1391–1397 (2015).
- S. Kapila, P. Nam, R. Mathur, D. Forciniti, S. Lorbert, and C. Schasteen, US Patent No. 6,605,590 B1, August 2003.
- S. Kapila and Monty Kerley, US Patent 8062681, November 22, 2011.
R. Seemamahannop, Radheshyam Panta, and S. Kapila are with the Department of Chemistry in the Missouri University of Science and Technology at the University of Missouri in Rolla, Missouri. Balaji Viswanathan is with Analytical R&D, Quality Control, at Pfizer Biotherapeutics in Saint Louis, Missouri. Direct correspondence to: kapilas@mst.edu















High-Speed Laser MS for Precise, Prep-Free Environmental Particle Tracking
April 21st 2025Scientists at Oak Ridge National Laboratory have demonstrated that a fast, laser-based mass spectrometry method—LA-ICP-TOF-MS—can accurately detect and identify airborne environmental particles, including toxic metal particles like ruthenium, without the need for complex sample preparation. The work offers a breakthrough in rapid, high-resolution analysis of environmental pollutants.


The Fundamental Role of Advanced Hyphenated Techniques in Lithium-Ion Battery Research
December 4th 2024Spectroscopy spoke with Uwe Karst, a full professor at the University of Münster in the Institute of Inorganic and Analytical Chemistry, to discuss his research on hyphenated analytical techniques in battery research.

Mass Spectrometry for Forensic Analysis: An Interview with Glen Jackson
November 27th 2024As part of “The Future of Forensic Analysis” content series, Spectroscopy sat down with Glen P. Jackson of West Virginia University to talk about the historical development of mass spectrometry in forensic analysis.

Detecting Cancer Biomarkers in Canines: An Interview with Landulfo Silveira Jr.
November 5th 2024Spectroscopy sat down with Landulfo Silveira Jr. of Universidade Anhembi Morumbi-UAM and Center for Innovation, Technology and Education-CITÉ (São Paulo, Brazil) to talk about his team’s latest research using Raman spectroscopy to detect biomarkers of cancer in canine sera.


